گیاهان سه کربنه C3 و چهار کربنه C4
گیاهان سه کربنه
تیپ گیاهان C3 که در اکثر گیاهان دیده می شود، به دلیل اینکه CO2 تثبیت شده در ساختار گیاه، در اولین مرحله از واکنشهای تاریکی فتوسنتز (چرخه کالوین) توسط آنزیم روبیسکو منجر به تولید قندی 3 کربنی موسوم 3- فسفو گلیسر آلدهید میشود، به این نام معروف است. این گروه از گیاهان با یک مشکل جدی مواجه هستند:
تنفس نوری
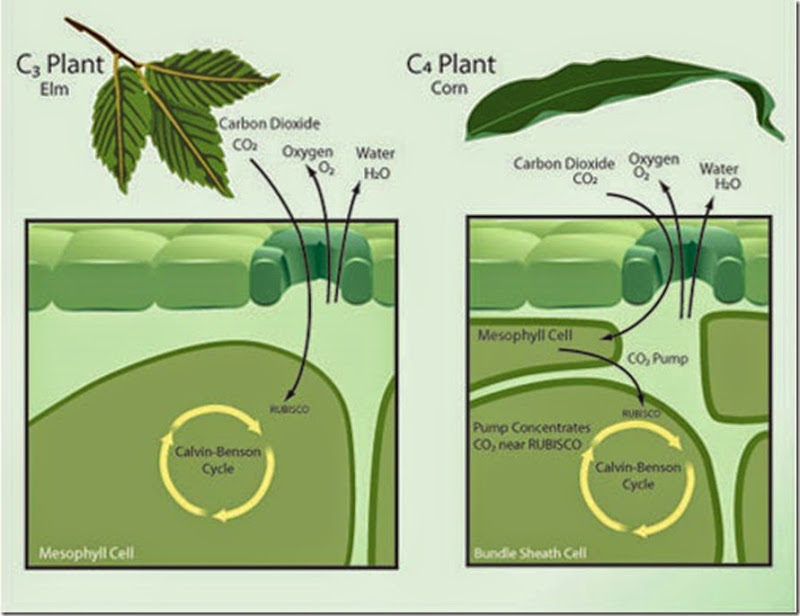
این فرآیند که به دلیل عملکرد دوگانه روبیسکو (کربوکسیلاسیون و اکسیژناسیون) رخ میدهد، منجر به کاهش بازده فتوسنتز در گیاهان میشود. دلیل این هم که این گیاهان بر خلاف گیاهان تیپ C4 قادر به حذف چنین فرآیندی نیستند به ساختار این دو دسته گیاهی برمیگردد: در گیاهان C4 محل انجام مرحله روشنایی فتوسنتز (مراحل مربوط به انتقال الکترون بین فتوسیستم I و II که منجر به رهاسازی اکسیژن به عنوان عامل آغازگر تنفس نوری میشود).
از محل انجام واکنشهای مرحله تاریکی (چرخه کالوین که روبیسکو در این چرخه قرار دارد) جدا است! به طوری که واکنش اول در سلولهای مزوفیلی و واکنش دوم در سلولهای غلاف آوندی رخ میدهد. این امر موجب می شود که اصولا اکسیژن آزاد شده طی واکنشهای روشنایی در مزوفیل به دلیل عدم دسترسی به روبیسکو به عنوان عامل تنفس نوری (که در درون سلولهای غلاف آوندی است)قادر به ایجاد تنفس نوری نباشد .
در طرف مقابل ، در گیاهان C3 چنین تمایز سلولی در انجام دو مرحله فتوسنتزی وجود نداشته و هر دو مرحله در سلول مزوفیلی رخ داده و اکسیژن آزاد شده طی واکنشهای نوری به عنوان سوبسترایی رقابتی با دی اکسید کربن بر روی جایگاه فعال آنزیم روبیسکو به رقابت پرداخته و منجر به وقوع تنفس نوری شود که این امر موجب کاهش بازده فتوسنتز در این گیاهان می شود . اصولا میزان تمایل روبیسکو به دی اکسید کربن خیلی بیشتر از تمایل آن به اکسیژن است. ولی به هر حال همین مقدار تنفس نوری هم میتواند منجر به کاهش بازده تنفس نوری شود.
در گياهان مختلف سبزينه دار كه فتوسنتز انجام مي شود و در نتيجه مواد غذايي ساخته مي شود اولين محصول پايدار بدست آمده در آنها متفاوت است . بدين معني كه اولين محصول پايدار در دسته اي از گياهان يك اسيد سه كربني به نام «3- فسفو گليسيريك اسيد» و دسته اي ديگر يك اسيد چهار كربني به نام« دي كربوكسيليك اسيد » ( داراي دو عامل كربوكسيل COOH- ، مثل اسيد اگزالواستيك ،اسيد ماليك ، اسيد آسپارتيك ) مي باشد . گياهاني كه اولين محصول پايدار حاصل از فتوسنتز آن ها يك اسيد سه كربنه است گياهان C3 و آندسته كه اولين محصول پايدار آن ها چهار كربني است گياهان C4 ناميده مي شوند . گياهان C4 در مقايسه با گياهان C3 از بازدهي فتوسنتزي بيشتري برخوردارند . بر همين پايه علف هاي هرز C4 نسبت به علف هاي هرز C3 قدرت رقابت زيادتري دارند . در تعدادي از گياهان گوشتي فرآيند فتوسنتزي ديگري مشاهده شده كه در شرايط رطوبت كم روزنه ها در شب باز شده و Co2 جذب مي كنند و در روز بسته مي شوند لذا شدت تعرق گياه خيلي كم مي شود . به اين نوع مكانيسم ، متابوليسم كراسولايي (CAM) “Crassulation asid metabolism” مي گويند مانند آگاو ،آناناس ،كاكتوس . كليه ي گياهان (CAM) جزءگياهان گوشتي غير نمكدوست هستند و عموما با محيط هاي خشك سازگارند .
برخي از گياهان C3 عبارتند از گندم ،جو ،سلمه ،ترشك ، توق ، تاتوره ،يولاف ،بارهنگ و پنيرك .
تمام گیاهان به انرژی نیاز دارند و این انرژی از طریق تنفس به دست می آید :
2 نوع تنفس وجود دارد :
1- مستقل از نور : قند و کربو هیدرات تولید شده در فتوسنتز را به CO2 و H2O تبدیل می کند. از نظر زراعی تنفس نشان دهنده تلفات است و باید به حداقل برسد که درست نیست زیرا تنفس لازم است تا انرژی برای رشد و بقای گیاه فراهم شود و تنفس بیش از نیاز برای تولید عملکرد زاید است . پس اهمیت سرعت اسیمیلاسیون خالص بیش از سرعت فتوسنتز و تنفس است.
2 – تنفسی که در حضور نور و در گیاهان C3 مشاهده می شود و ظاهرا در گیاهان C4 وجود ندارد.
گیاهان چهار کربنه یا C4 یا گیاهان کم CAM
گیاهان CAM گیاهانی هستند که در اثر شرایط سخت و خشک آب و هوایی به نوعی فرایند سوخت و ساز مجهز شدهاند که طی آن CO2 تنها در طول شب جذب گیاه میشود و در طول روز روزنههای هوایی گیاه برای جلوگیری از تبخیر اضافی آب بسته میشوند. CO2 جذب شده در طول شب به جوهر سیب (به انگلیسی: Malic acid) که یک ترکیب شیمیایی با شناسه پابکم ۵۲۵ است تبدیل میشود، همچنین آنزیمی به نام روبیسکو (Ribulose-1,5-bisphosphate carboxylase oxygenase) سبب افزایش بازدهی در تولید میشود.
تا اواسط دهه ۱۹۶۰ چنین تصور میشد که تمام گیاهان مسیر فتوسنتز ۳ کربنه (C3) دارند؛ ولی هاچ واسلک در سال ۱۹۶۶ ضمن مطالعه روی نیشکر متوجه شدند که اولین مادهای که پس از تثبیت Co2 در گیاه ساخته میشود یک اسید ۴ کربنه به نام اسید اگزالواستیک است. در گیاهان C4 هر دسته آوندی توسط یک لایه از سلولهای بزرگ پارانشیمی به نام غلاف آوندی احاطه شده است و این غلاف آوندی هم توسط سلولهای کوچکتر میانبرگ احاطه شدهاند.
به این ساختار برگ گیاهان C4 ساختار کرانژ (Kranz) یا غلاف آوندی میگویند. مسیر تثبیت Co2 در گیاهان C4 به این صورت است که در سلولهای میانبرگ این گیاهان ابتدا فسفوانول پیروات (PEP) با Co2 ترکیب شده و تولید اسید آلی چهار کربنه مانند اسید اگزالواستیک، اسید مالیک، اسید اسپارژیک میکند که این واکنش تحت تأثیر آنزیم PEP کربوکسیلاز صورت میگیرد. سپس این اسید ۴ کربنه تولید شده از طریق پلاسمودسماتاها به سلولهای غلاف آوندی منتقل میشود.
کم (CAM) به انگلیسی مخفف شدهٔ متابولیسم اسید کراسولایی است
در سلولهای غلاف آوندی این اسید ۴ کربنه Co2 خود را آزاد میکند و این Co2 وارد چرخه کالوین میشود و فرایند تثبیت کربن دی اکسید و تولید قند را موجب میشود. چرخه تثبیت Co2 از این مرحله به بعد مانند گیاهان C3 است. اسید ۴ کربنه که یک مولکول Co2 خود را از دست داده به پیروات تبدیل میشود که پیروات تولید شده به سلولهای مزوفیل برگشت داده میشود.
نوع دیگر تثبیت CO2 که CAM یا متابولیسم کراسولاسیون (Crassulation acid metabolism)، نامیده میشود بهطور عمده در گیاهان آبدار که دارای برگها یا ساقههای گوشتی هستند (نظیر آناناس – آگاو و کاکتوس) صورت میگیرد. چنین گیاهانی به شرایط خشک که کمی تعرق لازمه بقا است سازگار شدهاند. این دست از گیاهان در مناطق گرم و خشک، بیابانی، کمآب و مناطق حارهای امکان انجام فتوسنتز در روز را به دلیل تفات آب از طریق روزنههای خود ندارند بطوری که اگر روزنهها ی خود را در روز برای انجام فتوسنتز باز نگه دارند به دلیل تلفات شدید آب از روزنهها بصورت تعرق امکان حیات را از دست میدهند.
بنابراین در این گیاهان برای تطابق با شرایط بحرانی (از نظر دما و رطوبت) تکاملی در خود ایجاد کردهاند که در شرایط بسته بودن روزنهها در روز بتوانند فتوسنتز کرده و به حیات خود ادامه دهند.
مکانیسم تثبیت Co2 در گیاهان CAM شبیه گیاهان C4 است با این تفاوت که در گیاهان C4 محل فعالیت دو آنزیم PEP کربوکسیلاز و RUBP کربوکسیلاز متفاوت است ولی در گیاهان CAM زمان فعالیت این دو آنزیم فرق میکند.
گیاهان CAM در شب که دما و تعرق پائین است روزنههای خود را باز نموده و CO2 را بصورت اسید مالیک تثبیت کرده و در واکوئلهای خود ذخیره میکنند.
سپس در روز اسید مالیک ذخیره شده را به هیدراتهای کربن تبدیل میکنند (مانند گیاهان C4). به این ترتیب گیاهان CAM با انجام تعرق بسیار کم، فتوسنتز میکنند و به اینصورت به حیات خود ادامه میدهند.
اپیدرم گیاهان CAM از چند لایه تشکیل شده است و کوتیکول ضخیم دارند. سطح برگ آنها پوشیده از کرک است. در بعضی از گونهها کرکها کاملاً تحلیل رفته و بصورت خار درآمده است.
این گیاهان دارای برگها و ساقه گوشتی با واکوئلهای پر از آب هستند. گیاهان CAM مانند گیاهان CO2، C4 را بصورت اسیدهای چهارکربنه (توسط آنزیم PEPC) تثبیت میکنند و یکی ار تفاوتهای آن با گیاهان C4 اینست که این عمل در شب که روزنهها باز هستند انجام میشود. انرژی لازم برای این عمل از گلیکولیز تأمین میشود. در گیاهان CAM در طول روز مقدار هیدراتهای کربن ذخیرهای در برگ افزایش مییابد و در شب برعکس کاهش مییابد. تغییرات شبانهروزی در مقدار اسید آلی موجود در برگ گیاهان CAM بعلت تولید اسید مالیک در شب و تجزیه شدن آن در طول روز است. از جمله گیاهانی که دارای سیکل CAM هستند میتوان به انواع کاکتوسها، آناناس، آگاو و غیره اشاره کرد.
در شرایط رطوبت کم، روزنهها در شب باز شده و CO2 جذب میکنند و در روز بسته میشوند؛ لذا شدت تعرق گیاه خیلی کم میشود. گونههای دارای چرخهٔ CAM مانند گونههای C4، CO2 را بهصورت اسیدهای چهارکربنه با PEP کربوکسیلاز تثبیت میکنند و تفاوت آنها با گیاهان C4 در این است که این عمل در شب هنگامیکه روزنهها باز هستند درون واکوئلها انجام میگیرد. انرژی لازم برای این عمل را گلیکولیز تأمین مینماید. در روز تشعشع خورشید سبب بسته شدن شکاف روزنههای برگها میشود، لیکن این انرژی نوری جهت ادامه چرخهٔ کالوین در کلروپلاست بهکار میرود و CO2 از اسید چهارکربنه آزاد میگردد، نظیر آنچه که در سلولهای غلاف آوندی در گونههای C4 صورت میپذیرد. کلروپلاست گونههای CAM شباهت زیادی به کلروپلاست گونههای C3 دارد. فعالیت روزنهها و عمل کربوکسیلاسیون گونههای CAM در شرایط مرطوب و مساعد ممکن است بهصورت گونههای C3 تغییر پیدا میکند.
اختلافات گیاهان C3 و C4 :
1 – گیاهان C4 نسبت به C3 به نورهای با شدت بالا واکنش نشان می دهند.(2 برابر گیاهان C3)
2 – گیاهان C4 با کارایی بیشتری از CO2 استفاده می کنند و در شدت نور ثابت و نسبتا زیاد قادرند CO2 اطراف را به ppm 10 – 0 کاهش دهند. که این نقطه جبران است . در حالی که در C3 برابر
ppm 150 -50 co2 است . نقطه جبرانی پایین نشان دهنده کارایی زیاد فتوسنتز است .
3- گیاهان C4 سرعت اسیمیلاسیون خالص بالاتری از C3 دارند که به خاطر عدم وجود تنفس نوری است .
4 – درجه حرارت بر روی C4 تاثیر مطلوب ، و بر روی گیاهان C3 تاثیر نا مطلوبی دارد . تا دمای 0 تا 35 درجه سانتیگراد سرعت تنفس به ازای هر 10 درجه سانتیگراد افزایش حدود 2 تا 4 برابر افزایش می یابد. بسیاری از گیاهان C3 در محدوده دمایی 25 تا 35 درجه سانتیگراد تولیدی ندارند ولی گیاهان C4 افزایش نولید دارند.
5 – در گیاهان C4 کارایی مصرف آب بیش از گیاهان C3 است . میانگین ماده خشک تولید شده برای هر 1000 گرم آب مصرفی ، 29/3 گرم برای C4 و 54/1 گرم برای C3 می باشد.
6 – سرعت انتقال با سرعت فتوسنتز همبستگی دارد زیرا انتقال آهسته می تواند عملکرد را محدود کند . در برگهای گیاهان C4 انتقال 2 برابرسریعتر از برگهای C3 است.
7 – حداکثر رشد گیاهان C4 بیشتر از گیاهان C3 است (به جز چند استثناء). وقتی طول فصل رشد متوسط در نظر گرفته می شود سرعت رشد C3 برابر 13 گرم در متر مربع در روز بوده و برای گیاهان C4 برابر 22 گرم در متر مربع در روز می باشد. با محصور کردن گیاهان C3 و C4 در ظرف در بسته دارای نور اختلاف در تنفس نوری را می توان نشان داد . چون C4 نقطه جبرانی پایین تری از گیاهان C3 دارند زمانی که گیاهان C4 از CO2 مصرف می کنند گیاهان C3 از بین می روند.
گیاهان سه کربنه C3 و چهار کربنه C4
26
نوامبر